

WAGS NEWSLETTER
November, 2022
The WAGS Word
Welcome to the Q4 Western Ag Supply Newsletter!
Thank you for your business and we hope that as you read this, you and your families have been able to stay warm and have managed to dig out of the snowstorms we have seen around the prairies this week.
The last quarter has seen once again elevating grain prices with a lack of farmer selling but luckily, we have seen some strength in cash livestock markets at the same time. It sounds like feeders have largely pulled the trigger on purchases into the new year and are waiting to see if more sellers will come to market as the calendar year turns. The addition of another corn program in Southern Alberta shaping up could help to bring more grain supply to market which would be positive. In short, there should be some options as the calendar flips in terms of selection of ingredients to feed your livestock and our nutrition team will be ready to support you in those decisions.
Cue the segway to the introduction to the newest member of our swine nutrition team, Joaquin Sanchez Zannatta! Joaquin will be bolstering our knowledge and capacity as not only does he have several degrees focused on swine nutrition formulation and also spent a good portion of his career managing large sow operations. Joaquin looks forward to meeting many of you and has already gotten going working with some of our clients. Our nutrition group will continue to work as a team to make sure the requirements of all of our clients’ operations are satisfied.
WAGS is also very excited to announce the addition of David Raynard to our Saskatchewan sales team. David is originally from Swift Current and comes to us with a wide range of experience in sales, manufacturing, and business development. David will be a huge help in supporting our fast-growing Saskatchewan business.
On the warehouse operations side, we have the pleasure of Jake Adams joining our Montana operation. He has enjoyed getting to meet a good portion of our Montana customer base to date and looks forward to meeting more!
Welcome Joaquin, Jake and David!
Please enjoy this edition of the WAGS newsletter and we hope it provides you valuable insight on feeding and managing your operations.
Commodity Price Update
Grain and protein prices decreased over the past months following the good harvest progress across the Canadian prairies and the resumption of Black Sea shipments. However, prices have found support on the lack of farmer selling and strong domestic demand recently. On top of that, the uncertainties surrounding the export agreement between Russia and Ukraine added volatility to the market.
Although the Organization of the Petroleum Exporting Countries (OPEC) decision to cut crude oil production by 2 million barrels per day in November, the upside trend was capped by demand concerns due to global inflation and Covid-19 in China. As a result, Brent crude oil prices decreased from an average of $104 USD in July to around $98 USD in November. This trend could reduce production costs, but lower global diesel stocks may still provide upside risks. In addition, the uncertainties surrounding Russian exports could still drive prices upwards.
Bank of Canada has raised benchmark interest rate by 0.5 percentage points on October 26th. This moved the policy rate to 3.75% for the first time since early 2008. The Central Bank said that interest rates will likely need to rise further to get prices under control. The Canadian dollar is currently trading around $0.74 level. Despite Federal Reserve (FED) decision to hike interest rate by 0.75 percentage points, the Canadian dollar found support on good jobs data.
Corn prices received support from the strong US dollar, tighter than expected stock to use ratio in the US, and war uncertainties. Reduced production in Argentina and European Union are also bullish factors. Corn delivered to Saskatoon, SK is being traded around $455-$460 per MT and at $485 delivered to Red Deer, AB. La Niña could also reduce yields in South America, adding more volatility to the market. Notably, there is a corn import program shaping up starting sometime in December into the Lethbridge area with 7-10 unit trains of corn already booked for delivery Dec-April. Price delivered Lethbridge feeder is expected to be around $465/T.
Wheat prices are also increasing due to lack of farmer selling and strong domestic demand. Crop is achieving good quality grades, which could reduce the availability of wheat for feed. Delivered price indications in Red Deer, AB are at $465-$470 per MT delivered and around $470 per MT in Saskatoon, SK.
Strong feed demand and the bullish trend for grain prices have also driven barley prices upwards. Delivery prices in Lethbridge, AB are at $450-455 per MT Nov/Dec delivery and at around $400-$405 per MT in Saskatoon, SK. Due to the weakness of the Canadian dollar and good wheat crop quality across the Prairies, barley usage in feed rations has increased. Soybean meal pricing in Red Deer, AB is ranging around $700-710 per MT FOB Lethbridge. Prices have decreased following the US harvest, good crop perspectives in South America and weaker imports from China. However, prices found support recently due to concerns surrounding Black Sea shipments, cuts on US yields, and perspectives that China may increase its imports as crush margin improves. Following this trend, canola meal is being traded around $450-460 per MT in Red Deer, AB. Prices have decreased due to crush expansion and good Canadian harvest progress, and canola meal is providing strong value as a protein source in rations.
Pulse prices are around $465 per MT delivered Red Deer, AB and closer to $470-475 delivered Lethbridge area. Prices have increased due to strong demand, but still have good value in rations. Certain volatility is expected for grain prices as we move to Q4, especially because of the war in Ukraine. For proteins, prices could decrease following good crop perspectives in key producing countries, but Chinese demand will be an important driver.
Synthetic amino acids prices declined over the past months. This was led mainly by reduced freight costs and energy prices. Prices are expected to increase over Q4 following the Chinese New Year, which is a seasonal trend due to limited capacity of Chinese shipping ports.
IT IS A MOO POINT!
Ruminal metabolism of lipids and fatty acids
Lipids or fats are compounds that dissolve in organic (non-polar) solvents and insoluble in polar solvents such as water1. Therefore, in a feed analysis report, ‘crude fat’ may be shown as EE (ether extract or the compounds extracted by ether). A common word ‘lipids’ include solid fats and oils. Fat is a chemical combination of glycerol and fatty acids. A fat molecule consists of two components; a glycerol head (backbone) and three fatty acid tails (limbs)1. Glycerol is a small organic molecule with three hydroxyl (OH) groups, while a fatty acid consists of a long hydrocarbon chain attached to a carboxyl group (Figure 1). Typical fatty acids in feed contain 12–18 carbons, though some may have as few as 4 or as many as 361. Cows obtain fat from three primary sources; feed (grain and forages), fat supplements (etc. tallow, palm oil, fish oil, canola oil) and de novo synthesis of fat by microbes in the rumen and the cow’s body (eg: mammary gland)2. Table 1 shows fatty composition of fat supplements (canola oil and palm fat) common feed and cow’s milk.
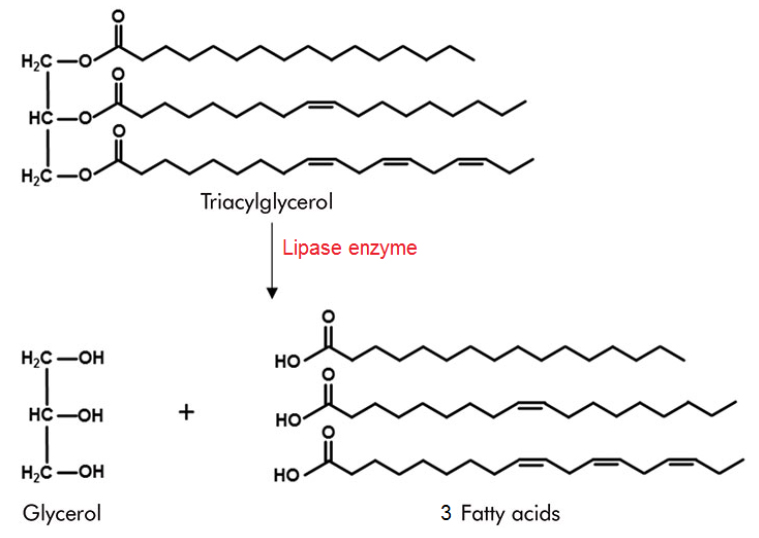
Figure 1. The process of lipolysis: Lipase enzyme acts a fat molecule and breaks it to 3 fatty acids and glycerol1.
Known fate of fat in the rumen: The changes happening to fat ingested by the cow, in the rumen is described commonly not as digestion of fat but bio-degradation6 and bio-hydrogenation2. Thereby a lipid sub-model is used to illustrate ruminal metabolism and intestinal digestion of long-chain fatty acids (LCFA). Ruminal lipolysis or fat digestion (defined as enzymatic cleavage of ester linkages and dissociation of salts of fatty acids) varies between feeds2. The enzyme that does the job is lipase and this rumen lipase would be from rumen microbes. Megalac (calcium salts of palm oil fatty acid distillate) was the only ingredient that showed appreciable protection against lipolysis and it would go to the intestines as by-pass fat.
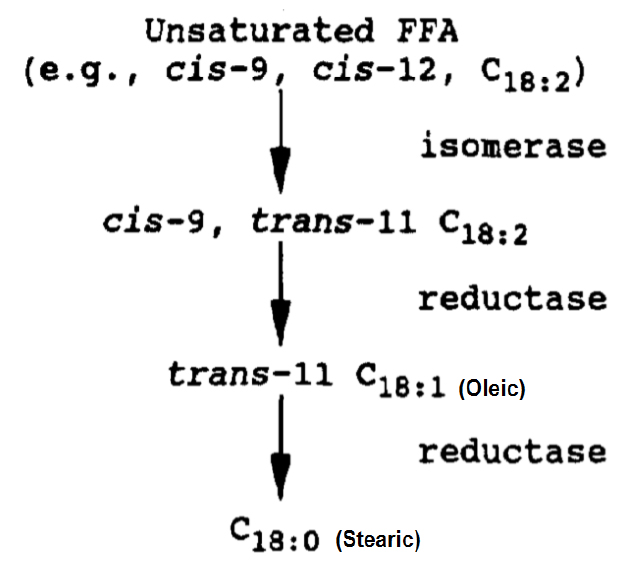
Figure 2. Bio-hydrogenation of fatty acids in ingested feed in the rumen by microbial enzymes (isomerases and reductases)6.
Lipolysis is a pre-requisite for ruminal bio-hydrogenation. Polyunsaturated fatty acids, especially linoleic (C18:2), appear to have an energy independent effect on improving cow’s reproduction. However, less than 10% of linolenic (C18:3), linoleic (C18:2) and palmitoleic (C16:1) that are in the form of free fatty acids can escape bio-hydrogenation (Figure 2). Oleic (C18:1), which can inhibit mammary synthesis of fat, accumulates in the rumen when there is incomplete bio-hydrogenation to stearic (C18:0, Figure 2).
De novo synthesis of fatty acids (production of extra fat) occurs (Figure 3), but as fatty acid intake increases, the extent of de novo synthesis declines as a result of the increased uptake of LCFA by microbes 2.
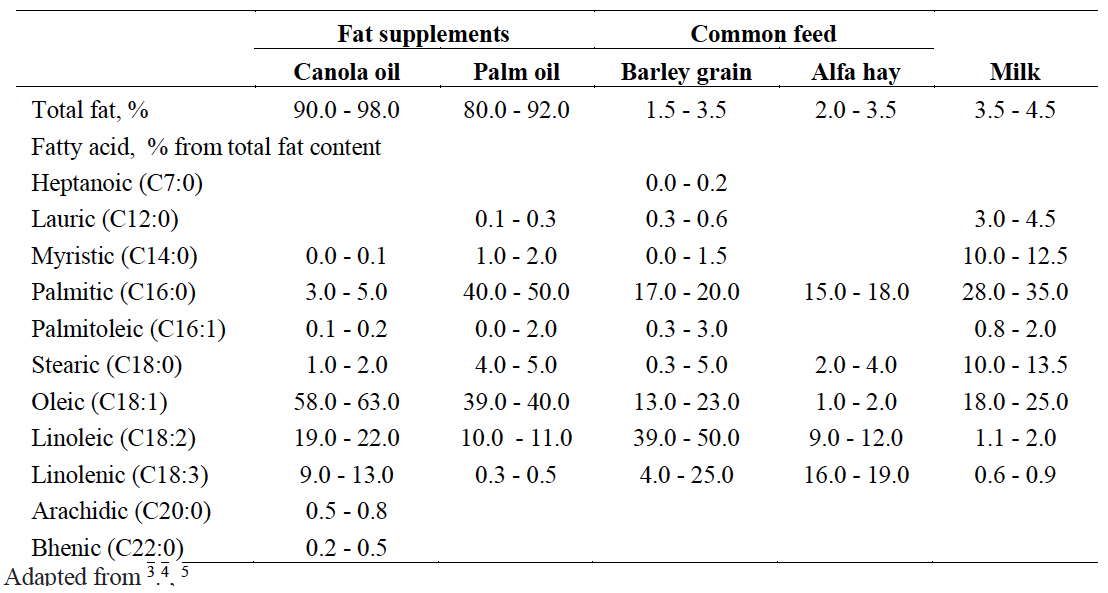
Table 1: Fat content and fatty acid composition in feed and bovine milk.
Data to model the effect of rumen active fat on rumen digestion and fermentation is limited7. However, the current recommendation of fat supplementation is based on the unsaturation of LCFA and ration fiber (forage to grain ratio), for the upper limit of total fat in the diet (Lass than 7% of TMR DM). In addition to the fiber level in TMR, breed and season of the year matter too8. Unsaturated fatty acids are more toxic to microbial cells than saturated fatty acids, and therefore, bio-hydrogenation in the rumen ecosystem is essentially a detoxification process. In other words, higher inclusion of certain fat types in TMR may exert detrimental effects on rumen microbes and deter the efficiency of rumen digestion. Understanding of ruminal fat metabolism focused primarily on manipulation of physicochemical events in the rumen aimed at two practical outcomes9:
1) Control of antimicrobial effects of fatty acids so that additional fat can be fed to ruminants without disruption of ruminal fermentation and digestion,
2) Regulation of microbial bio-hydrogenation to alter the absorption of selected fatty acids that might enhance performance or reduce saturation of meat and milk,
3) And, understanding and avoiding milk fat decrease caused by certain fat types 6.
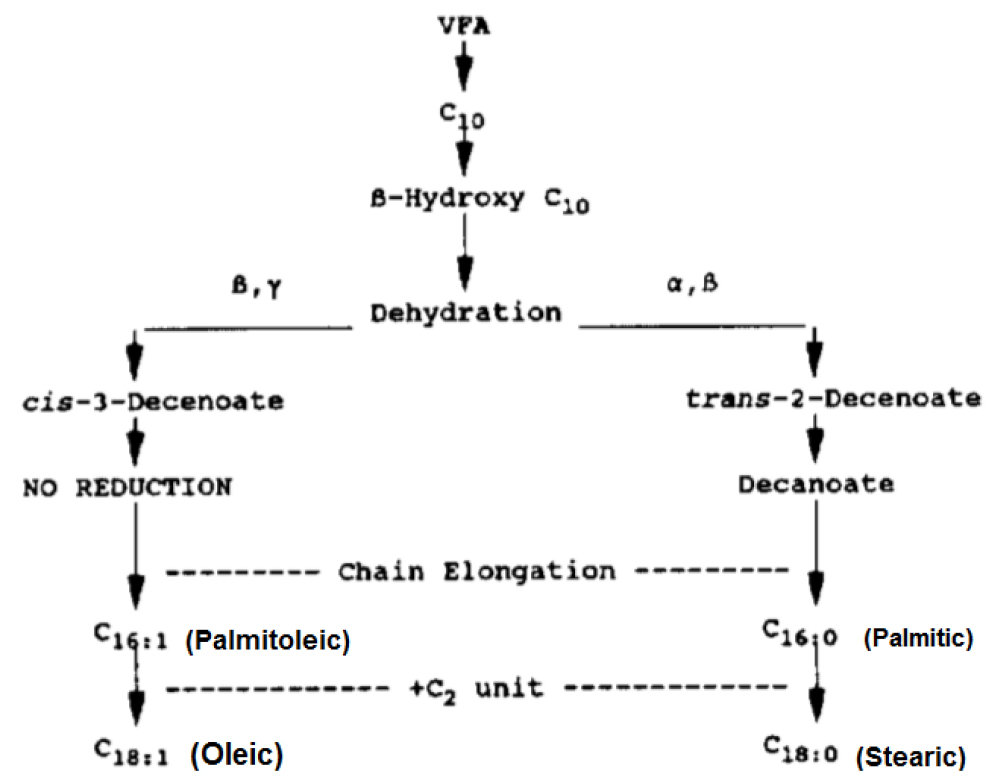
Figure 3. Fatty acids are synthesized by microbes in the rumen 6.
It is known that a better understanding of lipolytic and hydrogenation rates that vary with forage quality (stage of maturity and N content), surface area of feed particles in the rumen, and structural modifications of the fat molecule that inhibit attack by bacterial isomerases, to alleviate negative effects of fat inclusion and maximize the benefits 11.
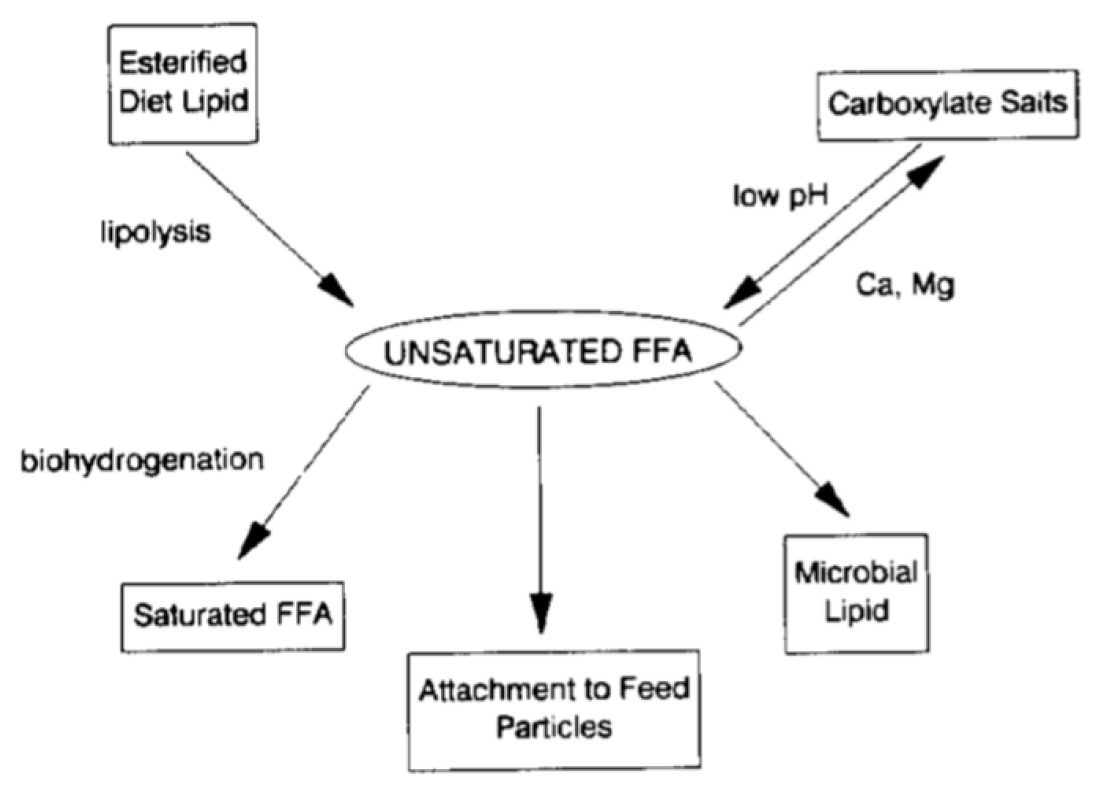
Figure 4: Free fatty acids undergo different changes caused by microbes inside the rumen 6.
Intestinal fat digestion: Similar to monogastric animals, fat coming to the small intestine, will undergo the digestion (lipolysis) by lipase enzyme (secreted by cow’s pancreas) activated at the presence of alkaline bile. Most of the LCFA flowing to the small intestine are free fatty acids but there are also non-lipolysed fatty acids in the form of glycerides and calcium salts and fatty acids in bacteria. In general, intestinal digestion of free fatty acids and non-lipolysed fatty acids in forages, grains, proteins, whole cottonseed, and cracked or ground soybeans and other oil-seeds are similar. Digestion of non-lipolysed fatty acids in tallow, hydrogenated tallow, grease, vegetable oils, animal/vegetable blends, whole-intact soybeans and other whole-intact oil-seeds with the exception of cottonseed is less than digestion of free fatty acids. In particular, digestion of non-lipolysed stearic (C18:0) in some feeds is zero. Digestion of non-lipolysed fatty acids in the form of calcium salts is greater than digestion of free fatty acids. To increase absorbed amounts of LCFA like linoleic (C18:2) and linolenic (C18:3), they must either be in a form that protects them from rumen lipolysis or an appropriate amount must be fed to achieve the target level, without disturbing the rumen. Nevertheless, it can lead to accumulation of oleic (C18:1) that can decrease mammary synthesis of fat causing decrease in milk fat.
Advancing knowledge: Many fundamental studies of ruminal fat metabolism motivated the practical uses of fat in cattle diets. One such motivation is the current interest to supplement fat to boost cows’ energy intake and body condition leading to healthy reproduction, without having detrimental effects of fatty acids on ruminal microbes and fermentation. Certain percentage amounts and types of fat supplementation may not be interfering ruminal fermentation 5.
However, further research studies on physicochemical effects of fat in the rumen are warranted. Key contributions are likely to come from detailed studies of how fat associates with surfaces of feed particles and microbial cells. The consequences of altered fat content and composition of microbial cells on ruminal digestive capacity are yet to be well understood. The motivation to increase absorption of unsaturated fatty acids continues because of their benefits as regulators of cell function, metabolism and about the possible health risks of saturated fatty acids in meat and milk for the consumers9. This dilemma is prone to get changed with ongoing research and meta-analysis of previous study data 12. Protected fat are needed to effectively hinder bio-hydrogenation without disturbing ruminal fermentation or altering intestinal fat absorption13. Research on control points of lipolysis and bio-hydrogenation may provide opportunities to discover positive and negative effectors of these microbial transformations 2. Therefore, feeding optimal fat content and optimal variation of fat types, both from common feed ingredients and fat supplements will be essential for rumen health, feed efficiency, body condition, energy levels, sound reproduction and milk fat content 14.
Note: References available upon request.
IT'S CHICKEN FEED
Biotin, vitamin D and vitamin E in poultry feeds
Vitamins are nutrients required by living creatures to carry out essential metabolic processes to keep them alive and healthy. Vitamins have highly specific functions; they are not synthesized by the host, are required in small amounts, and must be provided in the diet. The common food forms of the vitamins require a metabolic activation to become available in their functional form (Gerald et al., 2017).
Vitamins can be defined as organic compounds different than fats, carbohydrates, and proteins. They are natural components of feed ingredients, present in minute amounts which are insufficient to cover the requirements of the commercial modern chickens. In addition, those amounts are highly variable and cannot be taken into account when formulating diets and need to be added. When vitamins are absent or at low levels in the diet, deficiency signs are observed in the animals. Some of the functions the vitamins have in the body are related to hormones, antioxidants, enzyme, and cofactors (Gerald et al., 2017).
There are 13 groups of substances known as vitamins (Gerald et al., 2017). They are grouped into two major groups: water-soluble vitamins, and fat-soluble vitamins. Water-soluble vitamins dissolve in water and they are not stored in the body. Fat-soluble vitamins are absorbed along with fats in the diet and are stored in the body’s fatty tissues and in the liver. Vitamin A, D, E and K are fat soluble and Vitamin C, B1, B2, B6, B12, niacin, folic acid, biotin, and pantothenic acid are water-soluble vitamins. In this article we are going to discuss biotin, vitamin D and vitamin E. (Lykstad and Sharma, 2022)
Biotin. Biotin is also known as B7. It has a function as regulator in the gene expression and it also bounds to certain enzymes and activate them to carry their biological functions in the body. It plays an important role in the metabolism of fats, proteins, and energy (Gerald et al., 2017).
Biotin deficiency in diets for breeder flocks might cause low hatchability. In growing birds, biotin deficiency might result in reduced growth rate, high feed conversion, disturbed and broken feathering, dermatitis, and leg disorders. The dermatitis appears as dry scaly skin of the feet, then the skin of the foot pads and toes cracks leading to haemorrhages, scabs and infection. Similar lesions might be also found at the corners of the mouth and the vent. If the deficiency is prolonged and severe, lesions of the eyelids will develop (Whitehead, 1977).
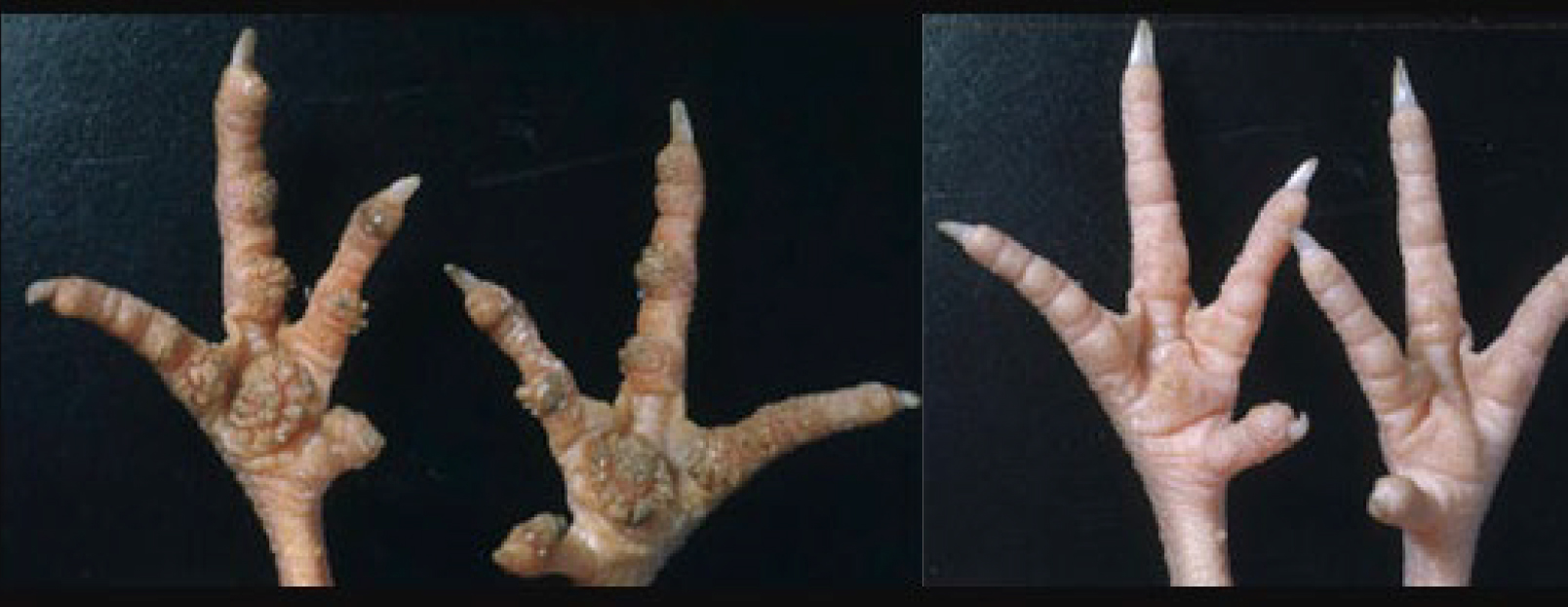
Fig. 5. The left-hand side shows a foot pad with dermatitis as a result of biotin deficiency and the right-hand side correspond to a normal foot pad.(Adapted from https://www-sciencedirect.com)
Vitamin D. Along with calcium, phosphorus and magnesium, vitamin D plays an important role in maintaining healthy bones. The forms of vitamin D added to the diets are not metabolically active, they must be converted in the kidneys and the liver to become functional (Gerald et al., 2017).
The active form of the vitamin D contributes to the absorption of calcium in the intestine, it also triggers the resorption of calcium and phosphorus in the kidneys, which allows the formation of the bones. In Vitamin D deficient scenarios, the normal absorption of calcium in the intestine is impeded and bone formation is affected causing rickets in growing animals and osteoporosis in grown animals (Gerald et al., 2017).
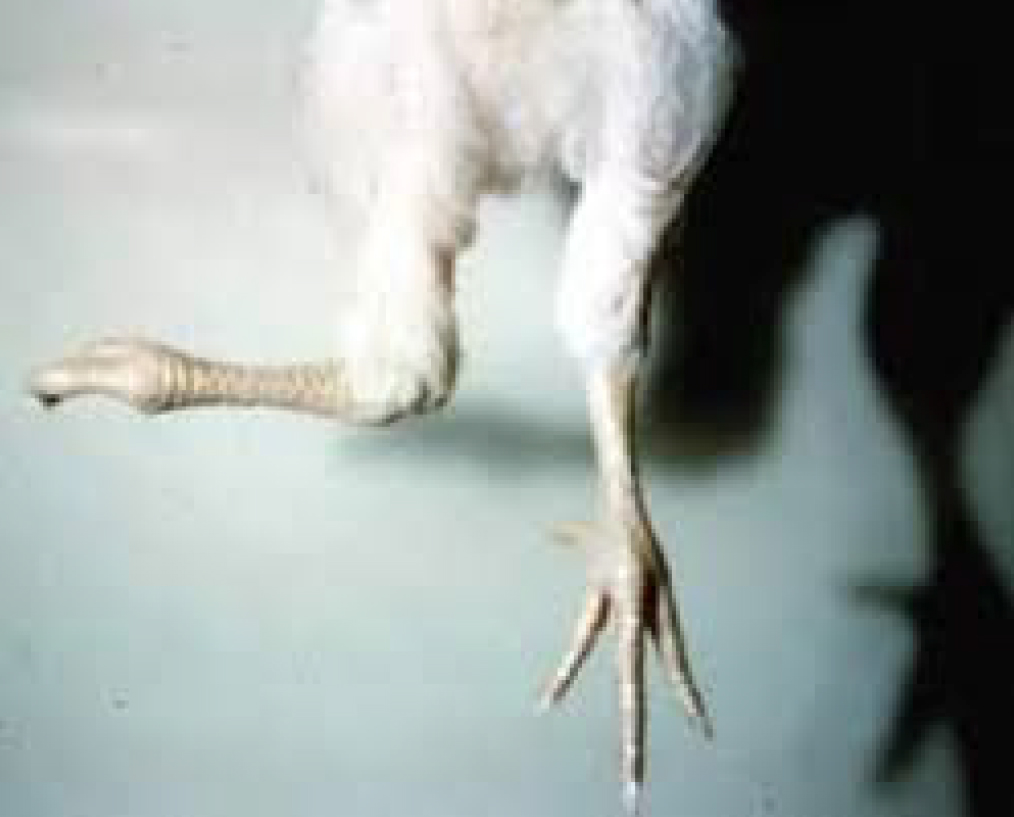
Fig. 6. Broiler with abnormal bone development or rickets. (Adapted from https://www.merckvetmanual.com)
In laying hen birds, bone strength and eggshell quality are related, this is because the calcium required to form the eggshell comes from the storage inside the bones, called the medullary bone, which is developed when the hens reach the sexual maturity. If there is a depletion of calcium in the medullary bone some of the calcium needed for eggshell formation may come from the structural part of the bones resulting in deterioration of bone structure, which may cause bone fractures and osteoporosis. Deterioration of shell quality may also be observed. Vitamin D enhances the absorption of calcium from the feed and consequently the formation of the bones; therefore, an adequate vitamin D supplementation is critical (Olgun and Aygun 2016).
Vitamin E. Vitamin E has a fundamental role as antioxidant in the metabolism of all cells. It protects the lipids from oxidation through the elimination of free radicals. Its deficiency can affect several organs. Free radicals are mainly produced in cells as by-products in the energy production process. They play an important role in physiological processes to keep the body healthy and properly functioning, however, they also cause damage to the DNA, proteins, and lipids, therefore vitamin E is important to keep the free radicals at normal levels to avoid the oxidation of the membrane cells whose main components are lipids. (Gerald et al., 2017).
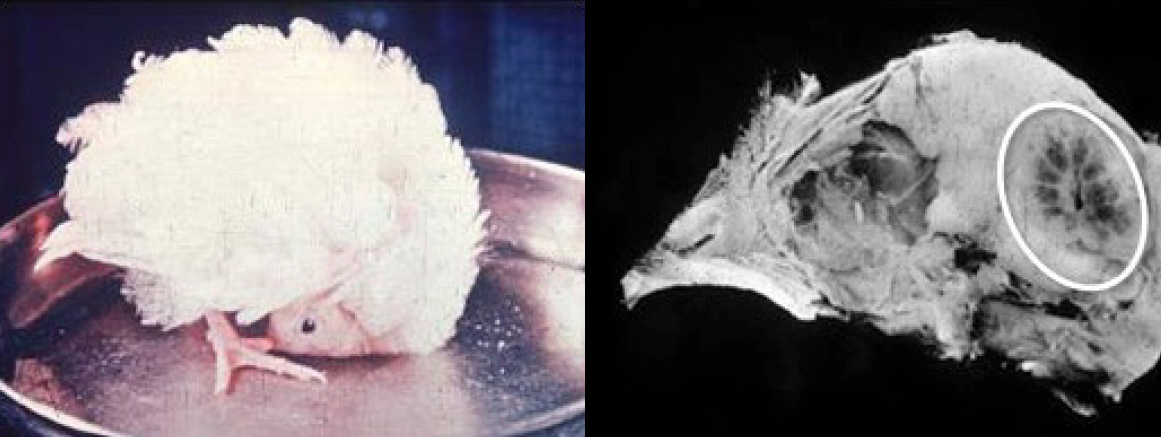
Fig. 7. Encephalomalacia in a vitamin E deficient chick. Post-mortem examination shows cerebral hemorrhage (Adapted from https://www-sciencedirect.com)
Nowadays, vitamins are manufactured and purified in commercial scale facilities and are available to the feed industry in very concentrated forms. Because of the high concentration careful dosage and mixing is required. Commercial premix manufacturers have the proper equipment and qualified personnel to provide feed manufacturers the required premixes containing the vitamins and minerals that are needed in the manufacturing of feeds (Leeson and Summers 2005). At Western Ag Supply we have access to world class suppliers that guarantee the use of best quality vitamins and minerals to provide our customers with superior quality micro-premixes and premixes.
Note: References available upon request.
PIG TALK
Piglet Creep Feeding
Piglets are subjected environmental and physiological stresses during weaning. Feeding solid feed or creep during nursing is a strategy commonly practiced to mitigate the negative effects of weaning. It is intended to supplement sows’ milk therefore, in theory, would result in heavier pigs at weaning, and since the pigs are accustomed to solid feed, should minimize post-weaning lag (Agyekum et al., 2017).
Does creep feeding improve weaning weights?
Several studies have shown that average weaning weights and average feed intake were not improved with creep feeding. However, this does not necessarily mean that creep feeding has no benefits. Data such as average weaning weights and average feed intake assumes that all piglets in the pen ate creep. It is estimated that the proportion of pigs that eat creep is only between 4 to 40% (Columbus et al, 2022; Seddon et al., 2016).
For example, in a study by Shea et al (2012), pigs offered creep in the crate 7 days before weaning, regardless of weaning age (d21 or 28), had the same weaning body weight as those not offered creep in the crate. However, pigs who were offered creep in the crates had higher nursery exit weights and daily feed intake compared to those who were not offered creep pre-weaning. Focusing only on the group that was offered creep feed in the crates, there were more pigs classified as “eaters” in pigs weaned at d28 (34%) than d21 (4%). The “eaters” had higher body weights at weaning and had higher growth rates in the nursery compared to “non-eaters”. These results suggest that creep feed is beneficial to pigs who consume it while in the crates and therefore efforts should be made to encourage piglets to eat creep prior to weaning.
Does the type of creep feeder matter? Seddon et al. (2016) offered creep feed in either standard feeder or in trays (Fig. 8). They observed that piglets had significantly more visits in the tray feeder than the standard creep feeder. Pigs offered creep feed in the tray feeder also had more “eaters” compared to those with standard feeder and had not post-weaning lag whereas piglets fed creep using a standard feeder did.

Fig. 8. Tray vs standard round creep feeder. (Photo courtesy of Prairie Swince Center Inc)
Some producers prefer to use creep feed in pellet or crumble form instead of mash and usually have preference for a small pellet size, 3mm or less. When Van der Brand (2014) offered piglet creep with larger pellet size (10 to 12 mm) or standard pellet size (2 to 3 mm), the piglets preferred creep with the larger diameter and feed intake was higher pre-weaning. In addition, pigs fed larger pellets had higher body weight gain and feed intake post-weaning and had no post-weaning growth lag. This has been validated in a multi-farm demonstration trial conducted in Canada (OMAFRA, 2016). The researchers in OMAFRA (Fig. 9) observed that piglets offered creep at 12 mm pellet had higher feed intake pre-weaning, had no post-weaning growth lag, and one farm had pigs that were 1.4 kg heavier at 28 days post weaning than those fed creep with a smaller pellet. It was explained that the large pellets also served as enrichment and encouraged the piglets’ exploratory behaviour and social interaction.

Fig. 9. Small vs large pellet size creep feed. (Photo courtesy of Dr. Laura Eastwood, OMAFRA)
Is it beneficial for the piglets to be on creep feed longer?
Studies suggest that feeding creep for longer duration does not improve pre-weaning performance. Sulabo et al (2010) fed creep feed for 13 (from d7), 6 (from d14) or 2 (from d18) days before weaning (d21) and showed no differences in weaning weight, ADG and ADFI. Feeding for 13 days however increased the number of “eaters” by 10% compared to feeding for 6 or 2 days.
Is a complex creep feed better than a simple diet?
A recent study by Columbus et al (2022) at the Prairie Swine Centre showed no difference in piglet weaning and post weaning performance when they are fed either a complex (with milk and other animal proteins) or simple diet (lactation feed; all plant-based) in the crates. When offered at the same time, piglets had no preference between simple or complex diet. The authors, emphasize that these data look at pen average and assume that all piglets eat creep feed in the crates. It has been previously established that only between 4 to 40% of the piglets eat creep feed and that those who do reap the benefits of creep feeding compared to those who do not eat creep feed.
The same group looked at nursery feed composition and found that diets with animal-based protein sources and amino acids methionine, threonine and tryptophan at 120% of NRC recommendation improved the piglets’ ability to fight off intestinal challenge such as Salmonella infection.
Does feeding creep also affect sow performance?
Milk output is improved by high density lactation feed intake and maximizing lactation feed intake and consequently, in theory should improve piglet growth rate pre-weaning (Sulabo et al, 2010). Pajor et al (2002) suggested that creep feeding augments sows’ milk in large litters and reduce the nutritional load on the sows especially during later stages of lactation, reducing the sows’ body weight loss and wean-to-estrus interval. In a study by Sulabo et al (2010), sows fed ad libitum lactation feed had 32% higher daily feed intake and 26% higher total feed intake compared to restricted-fed sows. Sows fed ad libitum had higher energy intake and thus resulted in lower body weight and backfat loss during lactation compared to restricted-fed sows. Interestingly, piglet growth rates and mortality rates were not different between the sows fed ad libitum or restricted-fed. A higher body weight and backfat loss was observed in the group of sows with restricted feed intake which is an indication of greater mobilization of body tissues. These observations suggest that if nutrient availability is limited, the sow will sacrifice herself to produce milk for her piglets.
There were no differences in the average days to estrus between ad libitum and restricted-fed sows, but the percentage of sows failing to return to estrus by d14 was higher in restricted-fed sows. Decreased feed intake during lactation suppresses the luteinizing hormone which is responsible for returning to estrus. It was suggested that feeding creep takes off some nutritional load from the sows. In this study, feeding the litters creep feed did not affect daily and total feed intake of sows. In addition, there were no differences in the BW at weaning or body weight loss during lactation, and proportion of sows who returned to estrus on d14. However, it was observed that creep feeding increased days to estrus among sows who returned to heat by d14. The authors did not offer an explanation.
Summary and conclusion: When, How and Where to feed?
The goal of offering creep in the farrowing crate is to create “eaters” at weaning which improves post-weaning pig performance by changing feeding behaviour. These are some of the ways to improve the creep intake of piglets on your farm:
When: Offer creep feed 7 to 4 days pre-weaning. Feeding creep for longer periods pre-weaning does not always increase piglet weaning weights but may increase the number of “eaters” which has huge benefits on piglet performance post weaning.
How: The decision to feed a simple or a complex creep diet lies on the farm challenges. There is evidence that piglets seem to have no preference between simple or complex diets and feeding a complex diet may not always improve piglet weaning and post-weaning performance. However, complex diets with animal-based protein can improve the piglet’s ability to fight off intestinal infections and may stimulate more piglets to eat creep.
Contrary to conventional wisdom, smaller pellet size (3mm below) is not necessarily better for piglet performance.
Where: The round standard creep feeder typically used in most farms can still be improved to stimulate creep feed intake. Feeder design affects feed accessibility and wastage. Trials have shown that tray feeders can help to increase the pig’s feeder visits and create more “eaters”, compared to pigs with standard feeders.
In conclusion, creep feed is beneficial to piglets who actually consume it and so efforts to increase the proportion of eaters in the litter prior to weaning should be made a priority to improve piglet performance in the nursery.
Note: References available upon request.
ANNOUNCEMENTS
INDEXING
1st New Dale Hutterian Brethren
2nd Wymark Farming Company Ltd.
7th Hillridge Farming Co.
8th Raley Colony Ltd.
10th Vanguard Farming Co. Ltd.
HEALTH/QUALITY TATTOO
1st Sand Lake Farming Co. Ltd.
3rd Vanguard Farming Co. Ltd.
7th R Valley Farming Co. Ltd.
8th Bench Farming Company
LEAN PERCENTAGE & LOIN DEPTH
Lean Percent target
1st Vanguard Farming Company Ltd.
2nd Wymark Farming Company Ltd.
3rd Bench Farming Company
8th Southland Farming Company Ltd.
9th New Dale Hutterian Brethren
Loin Depth Target
1st New Dale Hutterian Brethren
3rd R Valley Farming Co. Ltd.
10th Raley Colony Ltd.
Core Area and Sort
% in Core Area
1st Hillridge Farming Co.
2nd New Dale Hutterian Brethren
3rd Raley Colony Ltd.
Carcass Wt. Target
1st New Dale Hutterian Brethren
2nd Raley Colony Ltd.
6th Hillridge Farming Co.
7th Vanguard Farming Co. Ltd.
Congratulations and well done! WAGS is proud to be the nutrition provider in your barns.
JOKE TIME
A farmer notices that her neighbor produces a lot of big tomatoes every year, so she asks him what his secret was.
He says that he walks naked every morning as the sun rises.
The next harvest he checks up on his neighbor and asked if she had good tomatoes this year.
She says, “I walked around the garden naked every morning as you said but there were not that many tomatoes, but the cucumbers surely got big this year.”
(Source: jokojokes.com)
This little piggy
A pig walks into a bar and orders ten beers.
As soon as the pig is finished drinking the beers, he pays the bartender and starts to leave the bar.
"Wait!" says the bartender. "You drank so much beer. Wouldn't it be wise to use the bathroom before leaving?”
"Not for me," says the pig. "I'm the type of pig that goes wee wee wee all the way home."
(Source: jokojokes.com)
THE WAGS TEAM
David Borsboom – Sales Manager
Harry Korthuis – Sales Manager
Darcy MacDonald – Sales Manager
David Raynard – Sales Manager
Doug Richards – Sales Manager
Tony Rock – Sales Manager
Gordon Van Dasselaar – Sales Manager
Denni Van Dasselaar-Sales Manager
Anne Dyck-Office Assistant, Lethbridge, Alberta
Hailey Moors – Office Manager, Lethbridge, Alberta
Darlene Thorburn – Office Coordinator, Swift Current, Saskatchewan
Jake Adams– Warehouse Coordinator, Great Falls, Montana
Saman Abeysekara – Ruminant Nutritionist
Benjamin Londono – Poultry Nutritionist
Darren MacLeod – Swine Nutritionist
Mario Rebolledo – Poultry Nutritionist
Joaquin Sanchez Zannatta – Swine Nutritionist
Danilo Sotto – Swine Nutritionist
Tom Dowler – General Manager